家族罕见的端粒突变揭示了端粒灵活性和疾病的新见解

最近发表在《Nature Communications》杂志上的一项研究中,研究人员在端粒酶中发现了一个具有基因编码的端粒模板序列突变(TTAGT)的家族,这导致了一个非常规端粒序列至少延续了一代。
研究报告说,端粒序列 TTTAGG 在所有脊椎动物中都是恒定的,并通过与一组称为 shelterin 的蛋白质结合,在抑制脱氧核糖核酸(DNA)损伤反应方面发挥着关键作用。端粒序列的变化会减少 shelterin 结合,造成 DNA 损伤,并对细胞有毒。

关于研究
在本研究中,研究人员发现了一种新型的杂合突变,它是一种对端粒序列编码的特发性肺纤维化患者的 TERC。这种变异使患者能够在四十多年内保持无病状态,并被遗传下来。
最近,研究人员分析了特发性肺纤维化病毒的患者,发现了一些罕见的 TERC 和 TERT 突变。一个个体在 TR 模板区显示杂合子型 C>A 翻转。病史包括头发过早变白,43 岁早期诊断为特发性肺纤维化(IPF)。为了验证体内含有 r.50C>A 基因变异的端粒酶活性,研究人员分析了先证者及其儿子的全基因组测序(WGS)结果,以研究变异序列与端粒的整合。
研究人员利用外周血单核细胞(来自原细胞的PBMC)或唾液(来自子细胞的)样本来计算野生重复和突变体在原细胞、子细胞和十个对照中的百分比。他们使用定制的 Perl 程序计算野生类型和变异重复。接下来,他们用 PNA 探针对三个序列变异或野生型重复进行了现场变异端粒序列识别。研究中的原体进行了肺移植,并利用荧光原位杂交(FISE)对野生型端粒和突变进行了研究。
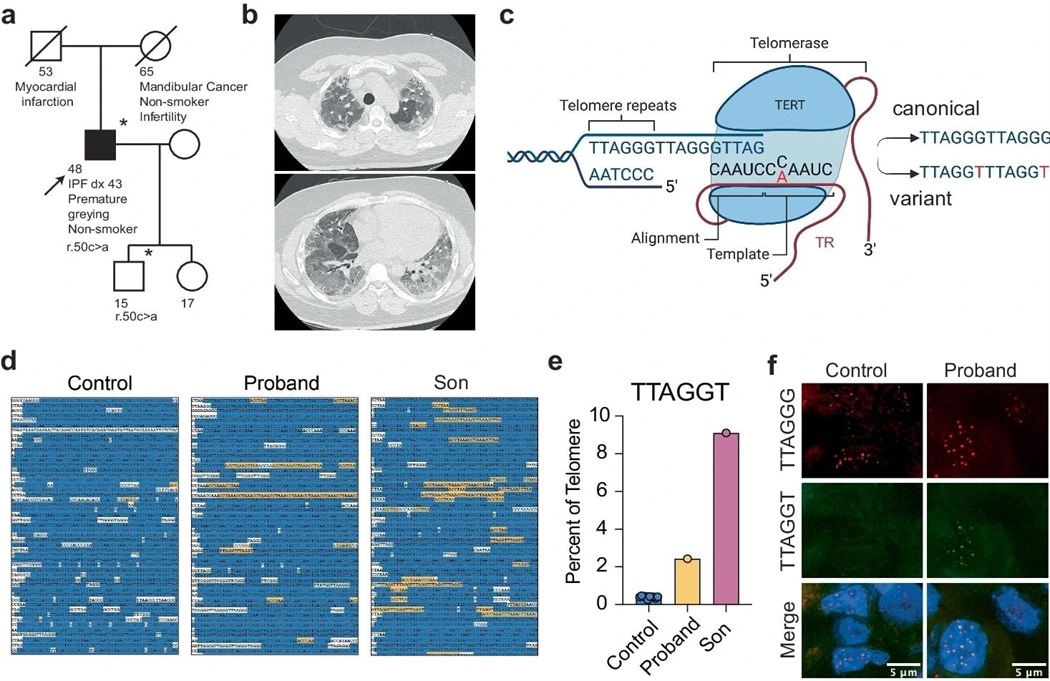
a. 携带 TERC r. 50c > a变种的家庭系谱。先证者(箭头)在43岁时被诊断为IPF。星号表示可获得DNA的个体。正方形表示男性,圆圈表示女性。符号上的一条线表示该人已经死亡。b. 在肺移植前,胸尖(上)和基底(下)CT显示原体间质改变和晚期纤维化。c. 图形显示患者派生变异在TR模板和结果端粒序列的位置。这个数字是在生物资源网站上创建的,并在一个共享的"非商业的"无衍生物第4.0国际许可证下发布的。d. 从一个没有模板突变的个体的整个基因组测序中提取的端粒体序列,原原体和原原体的子细胞。标准重复在蓝色中被突出,变异的Ttagt重复在黄色中被突出,额外的非标准序列在白色中。e. 在控制器中重复的TTAGT的百分比,主犯和主犯的子细胞。f. 野生型(红色)和变异型(绿色)序列在原移植肺和对照捐赠肺的组织切片中的应用。(e)的源数据作为源数据文件提供。
研究人员为变异的 TTTAGT 序列建立了表达构造,由于 R.C56 的 TR 与突变端粒的 3'端不匹配,可能导致重复加成过程(PB)不足。他们还研究了 POT1-TPP1(一种公认的RAP促进剂)是否可能在与 C50A 相互作用时恢复较差的 RAP。
研究人员研究了将变异序列引入 TERT 阳性细胞系,用编码多种变异的慢病毒转导细胞,以及用肽核酸(PNA)-FISH 探针检测端粒掺入的动力学和效果。他们还测量了研究先证者的端粒长度,并分析了端粒酶变异对3'端粒末端序列的影响。他们还研究了 POT1 是否可能在体外结合突变序列并阻断 POT1 介导的端粒酶活性抑制。
研究结果
分析表明,端粒酶基因的突变携带了一代或几代人,尽管几乎 9.0% 的端粒改变为新的序列,但一位家庭成员报告了没有重大的医疗问题。突变模板抑制端粒酶重复添加过程性和减少 POT1 相互作用。尽管有这些异常,这个序列很容易被整合到细胞染色体中。变异序列的存在可能会影响端粒的添加。
虽然他报告说没有严重的医疗问题,但是他的孩子也有这种变异。研究人员预计,该突变 R.50C>A 将引入 TTAGT 非常规型端粒基因序列,而不是 TTAGG。原带端粒的 TTTAGT 序列为2.7%,而对照组的 TTTAGT 序列为 0.40%(原带的序列比前),原带子的变异序列为 9.20%,增加了 23.0 倍,表明端粒酶对生殖系或发育过程有显著的重复添加。
先证者有恒定的规范性重复(84%),但他的孩子比对照组少(78%)。大约 15% 的端粒组成部分既不包括 TTTAGT 序列,也不包括野生型序列,这与以前使用 WGS 数据研究端粒构成的研究相类似。移植的肺组织包括可变的端粒序列,但对照供体肺没有。测序结果和原位杂交表明端粒酶吸收了成人组织中残留的序列。
结构模型表明,dG:rC 配对在拉链上比 dT:rA 配对更稳定。在 hTERT-RPE 细胞中,36% 的 C50A- 和 70% 的 c50 / 56a -转导细胞表现出至少 10 个突变的端粒灶,连续 TTAGGT 重复序列减少。先证者及其子女遗传变异 TR 的可能性分别为 39% 和 28%。
研究发现端粒可以承受相当大的退化,这意味着插入一个非常规端粒序列可能会影响端粒长度动态。由 C50A TR 组成的端粒酶由于突变体端粒和 TR.56C 之间的不匹配而导致实际上的全价损失。通过产生 C50/56A 变异来纠正错配,增强了区域行动方案,促进了更多的 POT1 - TPP1 相关的激活。然而,当过度表达时,C50/56A TR 与野生型 TR 一样延长端粒。突变的端粒序列,如 POT1ΔOB,可能会阻止保护素抑制端粒酶的添加。
期刊参考
Hinchie, A.M., Sanford, S.L., Loughridge, K.E., et al. A persistent variant telomere sequence in a human pedigree. Nat Commun 15, 4681 (2024). DOI: 10.1038/s41467-024-49072-9, https://www.nature.com/articles/s41467-024-49072-9